























The Humans We Haven’t Met Yet

Please note that this article includes images of human remains.
Everything that’s been called Homo sapiens, isn’t.
In my opinion, far too many species have been lumped together into this one taxonomic category. The truth of the human story is far more complicated, with more species and even more genera than have been named, and more dead ends on the branches of the human family tree than have been recognized.
As an evolutionary biologist, I’ve always been interested in the theory and practice of how one defines a species and tries to figure out its evolutionary relationships. I came to understand that this process is only ever hypothetical, never provable.
I began in the 1970s studying early mammals, then lemurs and other prosimians, then apes. I stayed away from hominins at first: There was too much storytelling and not enough evidence for my tastes. But my frustration with how the human fossil record was interpreted pushed me into this arena in 1997. My first goal was to put together an encyclopedia of as many human fossils as I could get permission to study. This led to three volumes, the last published in 2005.
Since then, I’ve continued to document the human fossil record with detailed descriptions and lots of photographs. And the more specimens I study, the more I realize that most of the species designations don’t make sense. If hominin fossils were treated the same way as nonhuman primates, specimens currently lumped into the same group would be allocated to different ones. We need to go back to taxonomic basics.
Having studied so many human fossils is both a blessing and a curse. The “blessing” is that I can apply my theoretical and comparative background to virtually the entire human fossil record. The “curse”: I don’t always see eye to eye with other paleoanthropologists, who often focus on one or two “species,” a geographic area, or a period of time.
We humans are the only surviving species of our immediate evolutionary group. This often leads people, including researchers, to make an intuitive but not necessarily correct assumption: The closer one gets to the present, the fewer contemporary species there should be.
To track how this came to be, we could start in 1735, when Carl von Linné—commonly called Linnaeus—published Systema Naturae. There he classified each plant and animal he knew of using a new strategy: the couplet of genus plus species. He also insisted that “genus” and its subordinate “species” be defined or diagnosed through specific morphological or behavioral features and not, as was common, by some “feeling.”
Yet, when it came to H. sapiens, Linnaeus ignored his own requirement. Instead of providing a morphological diagnosis of H. sapiens, he let us define ourselves with the phrase Nosce te ipsum (know thyself). This notion of “I know it when I see it” still informs today how hominin fossils are allocated to our species, H. sapiens.
Although rock-hard objects with the shapes of bones and teeth had been unearthed for centuries, only in the 1600s did Nicolas Steno, a Danish scientist, convince skeptics these were the fossilized remains of extinct animals. For western Europeans, this was a big deal. In the eyes of stalwart Creationists, God would not have allowed animals to go extinct. While the first human-like fossils were discovered in 1829, it took decades before their possible antiquity was even discussed.
The first publicly debated human-like fossils were discovered in 1856 at Feldhofer Grotto in Germany’s Neander Thal (German for valley). When Creationist and anatomist Hermann Schaaffhausen described these bones in 1857, he argued they were the fossilized remains of some brutish race that preceded living humans. In 1864, the geologist William King created a new species name for them: Homo neanderthalensis.
The late 19th century and early decades of the 20th century witnessed a rash of discoveries of now-accepted human-like fossils. This included more Neanderthals, Cro-Magnon and other Upper Paleolithic H. sapiens, along with specimens for whom their discoverers created new species, and even new genera. Indeed, from 1864 to 1949, paleoanthropologists named at least nine human-like genera and 12 species (see image below).
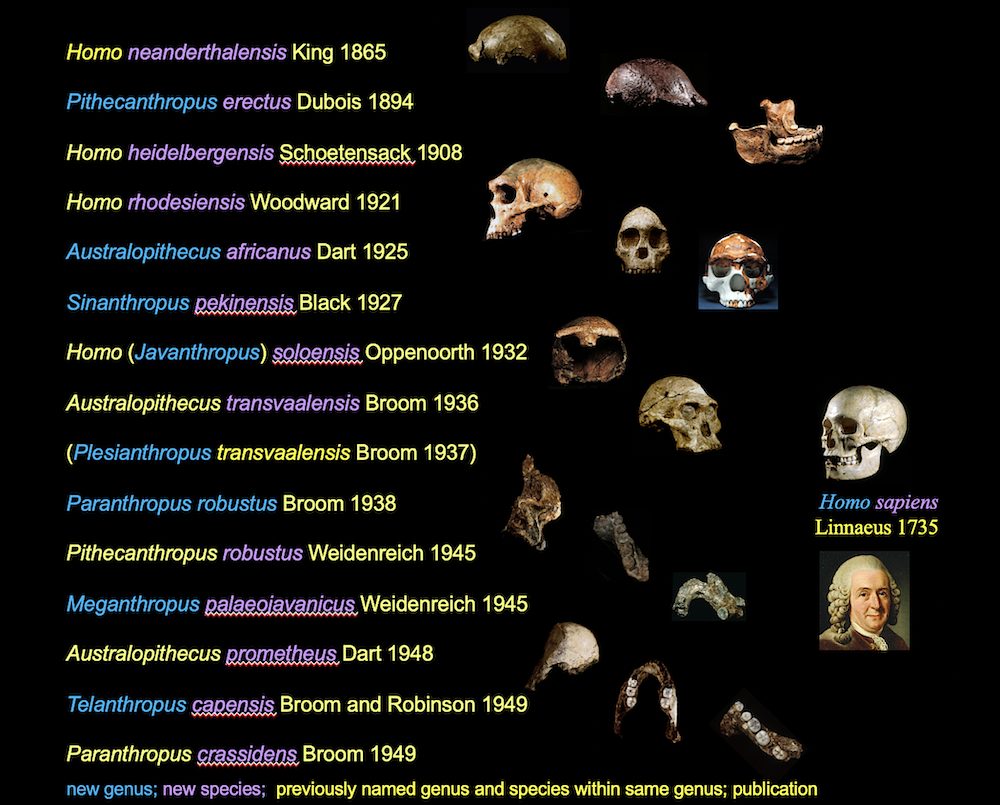
As early as 1933, physical anthropologist Solly Zuckerman argued that Neanderthals, Pithecanthropus (“Java Man”), Sinanthropus (“Peking Man”), and a skull from Zambia dubbed Homo rhodesiensis were morphologically so different from us and Upper Paleolithic H. sapiens, they didn’t belong in the genus Homo. In 1940, he suggested that this broader Neanderthal-like group deserved its own genus name.
So, while paleoanthropologists were latecomers to fossil hunting, in the 1800s and early 1900s, the emerging picture was that human evolution was taxonomically diverse and species rich. Just like other animals.
But this picture began to change in the 1940s.
In 1944, with World War II raging, pioneering Ukrainian geneticist Theodosius Dobzhansky argued that humans evolved in a different way from other animals because of culture: Instead of different species emerging as humans adapted and evolved to suit different particular circumstances, culture allowed humans to alter their environment to suit them. Thus, while very early on in evolution human populations may have begun to speciate, culture stopped the process. Therefore, he argued, the picture of human evolution was not one of diverse species, but of a single, variable lineage that changed over time. His argument was, in part, a critical response to the Nazi belief in “inferior races.”
Read more, from the archives: “Were Neanderthals More Than Cousins to Homo Sapiens?”
In 1950, taxonomist Ernst Mayr formalized Dobzhansky’s “no speciation” notion by collapsing all human fossils into genus Homo and three chronologically defined, transitioning species: transvaalensis (oldest), erectus (middle), and sapiens (youngest). That was all. They were thought to have marched from one species to the next in a chronologically simple line.
In doing so, Mayr demoted previously noticed differences between fossils to just individual variation within species. H. sapiens now included us, Upper Paleolithic humans, Neanderthals, and specimens previously attributed to H. rhodesiensis, Homo heidelbergensis, and Palaeanthropus palestinus. Echoing Dobzhansky, the underlying message was: If all of these morphologically disparate hominins were members of the same species, how could anyone think that differences between living human “races” were evolutionarily meaningful?
In 1963, Mayr recanted a bit, and “allowed” the “transvaalensis” specimens to be returned to their original genera. (It was now OK for there to have been some taxonomic diversity early on in human evolution.) But the rest remained the same. Paleoanthropologists still plunked specimens into erectus and sapiens primarily because of their age, not their morphology.
And as the number of morphologically disparate specimens in each “species” grew, so, too, did wonderment at how incredibly variable each species was.
In order to acknowledge the marked differences between human samples, paleoanthropologists in the 1960s began divvying H. sapiens into “archaic” and “anatomically modern” groups.
“Archaic” subsumed Neanderthals, with their low-rising, often long braincases, rounded brows, and large, puffy, projecting snouts. It included H. heidelbergensis with its robust lower jaw. And, like Zambian H. rhodesiensis, it included skulls with sloping profiles, thick brows, and chunky faces.
“Anatomically modern” humans were chronologically younger, and included us, Upper Paleolithic humans, and specimens from various Levantine and African sites with slightly rounded skulls, slightly bulbous frontal bones, less robust brow ridges, and, sometimes, smaller faces.
By 1990, H. neanderthalensis was firmly reinstated as a unique species. Not long after, most paleoanthropologists agreed that the other “archaic” specimens should be removed from H. sapiens and placed in H. heidelbergensis.
Even absent these specimens, H. sapiens was still a motley lot. Take, for example, the Skhūl V individual.
Skhūl V and many specimens still classified as Homo sapiens are about as different from us as Neanderthals are.
This specimen, dating to around 101,000–81,000 years ago from Skhūl Cave, Israel, is part of a group of skeletons excavated in the late 1920s to early 1930s that were named Palaeanthropus palestinus. Because of Skhūl V’s protruding face and prominent brow, anthropologists of the time thought its species bridged a gap between Neanderthals and Australian “Aborigines.” After 1950, the Skhūl specimens, like so many others, were lumped into H. sapiens.
Yet, in addition to its prominent brow and protruding face, in 2015, researchers pointed out that Skhūl V’s long, shallow palate would not have housed a short, thick tongue like ours, and, based on aligning the neck vertebrae, its voice box lay behind, not above, its breastbone. In short, this individual didn’t look like us, and it couldn’t produce the basic sounds of human speech.
Why is Skhūl V still considered a member of H. sapiens? It beats me.
Skhūl V and many specimens still classified as H. sapiens are about as different from us as Neanderthals are. Having studied them all, it seems the only thing we all have in common is that we don’t have long braincases with sloping foreheads.
From my decadeslong study of recent human skulls from all continents, I’d say H. sapiens has a few key physical features. There is no continuous brow above the orbital sockets and across the nasal region. Our lower face is vertical and a lot narrower across than our cheekbones; you can feel this yourself. Most importantly, we’re the only living mammal with a true chin: not just a bulge but an upside-down T shape that often becomes triangular as a person grows older.
Using such features to define H. sapiens, many fossils, including Skhūl V, as well as Omo 1 and II and those from Herto, Ethiopia, don’t make the cut. All European Upper Paleolithic humans, on the other hand, such as from Cro-Magnon, do have all of these features, so they’re H. sapiens. I would say the oldest collected specimen of H. sapiens is from Border Cave, South Africa, which is perhaps as old as 174,000 years.
In my view, the pot currently labeled H. sapiens contains specimens representing at least a few species, and these species are our closest extinct relatives. Even closer than Neanderthals and the recently described “Dragon Man,” proposed as Homo longi, from China.
Many people think that DNA accurately reveals the picture of human evolution. But this isn’t true.
First of all, DNA has only been successfully extracted from a handful of fossils, including some Neanderthals, some Upper Paleolithic and near-recent humans, a partial finger bone from Denisova Cave, and two bones from Sima de los Huesos, Spain. Since DNA usually degrades within 100,000 years, we’ll only ever have DNA from a minuscule fraction of fossils; DNA analyses are missing most of the pieces of the puzzle. And there are circular arguments: DNA is identified as, say, Neanderthal because the fossil was identified as such, and then it is used to identify other fossils as Neanderthal, even if they don’t look Neanderthal-like.
While certain similarities between our DNA and Neanderthal DNA have been interpreted as evidence of the two species interbreeding, these similarities could just be run-of-the-mill features of the genome, common to many species.

In the crush to embrace interbreeding as the source of human-Neanderthal molecular similarity, people sometimes forget to ask the question: If humans and Neanderthals actually crossed paths as often as molecular anthropologists claim—which is not supported by archaeological or paleontological evidence—would they have recognized each other as potential mates? I don’t think so: They looked too different. Even present-day hunter-gatherer groups have been observed to engage in violence when they encounter each other. And there is evidence from the 49,000-year-old site El Sidrón, that Neanderthals cannabilized other Neanderthals.
When Dobzhansky first kicked back against a taxonomically diverse picture of humanity, his ideas conveyed the well-intended implication that, if specimens as wildly different as Neanderthals, H. rhodesiensis, and modern humans all belong to the same species, differences between living humans become insignificant. We are all one, absolutely equal, no lesser group among us. This is true, of course: All modern humans are clearly, morphologically H. sapiens. But our present world should not cloud our view of the past.
While Dobzhansky’s reaction to Nazi ethnic cleansing was commendable, the legacy he left, which Mayr translated taxonomically, still constrains how many paleoanthropologists view the later phases of human evolution: More linear than species rich.
To me, however, all of human evolution, even well after our own species first emerged, is a mess of branches and dead ends. Our evolutionary history is complicated, and we should embrace that.

Jeffrey H. Schwartz is an emeritus professor in the departments of anthropology and history and philosophy of science at the University of Pittsburgh, and a research associate in the division of anthropology at the American Museum of Natural History. His publications span evolutionary and systematic theory, primate (including human) evolution and systematics, developmental and theoretical biology, and bioarchaeology. Schwartz’s books include The Human Fossil Record series, Sudden Origins: Fossils, Genes, and the Emergence of Species, The Red Ape: Orangutans and Human Origins, and Skeleton Keys: An Introduction to Human Skeletal Morphology, Development, and Analysis. In 1997, he began a detailed study of the human fossil record that has now encompassed virtually all known skulls, jaws, and teeth, as well as numerous pelvic and leg bones. Schwartz has been invited to write critical reviews of the uses and abuses of DNA analyses in human evolutionary studies, the most recent on mitochondrial DNA.
